
Einblicke in die komplexe Dynamik der Stickstoffumsätze im Boden
Die Stickstoffdynamik im Boden ist ein komplexes System, das durch verschiedene Faktoren gesteuert wird. Dennoch ist klar, dass Bodenbearbeitungen keineswegs immer vorteilhaft sind. Generell sollte auf eine Bodenbearbeitung in den Wintermonaten verzichtet werden. Bilder wie das obige sollte es im Winter besser nicht mehr geben.

In natürlichen Ökosystemen bildet der Stickstoff (N), der in der organischen Bodensubstanz (OBS) oder im «Humus» gespeichert ist, die Grundlage für die Versorgung der Pflanzen. Aus diesem Vorrat wird N mit Hilfe von Mikroorganismen mineralisiert und in pflanzenverfügbare Formen wie Nitrat oder Ammonium und zu einem kleineren Teil in organische Makromoleküle wie Aminosäuren überführt, die von den Pflanzen aufgenommen werden können.
Die Mineralisierungsprozesse sind im gemässigten Klima Europas sehr stark vom Jahresgang der (Boden-)Temperatur und der Bodenfeuchte abhängig. So werden z.B. auf leichten sandigen Böden mit geringeren Humusgehalten 40–115 kg N/ha im Jahr mineralisiert, auf schweren Böden dagegen nur 30–75 kg/ha (Tab.).
 Tab.: Jährliches Stickstoff-Mineralisierungspotenzial unterschiedlicher Weinbergsböden. (Ergebnisse aus 1880 Bodenanalysen, Rheinland-Pfalz, Deutschland, 2012)
Tab.: Jährliches Stickstoff-Mineralisierungspotenzial unterschiedlicher Weinbergsböden. (Ergebnisse aus 1880 Bodenanalysen, Rheinland-Pfalz, Deutschland, 2012)
Rezirkulierung und Rhizodeposition
Der von den Pflanzen aufgenommene N wird wiederum im N-Kreislauf rezirkuliert. Dabei spielen Einträge über die Wurzeln und oberirdischen Rückstände der Pflanzen eine grosse Rolle. Lebende Wurzeln geben sowohl Kohlenstoff (C) als auch N-Verbindungen wie Wurzelexudate, Mucigel, Wurzelhaare oder abgebaute Feinwurzeln in den Boden ab. Diese Einträge werden als Rhizodeposition zusammengefasst. Über Rhizodeposition können erhebliche Mengen N in den Boden eingetragen werden: Sie betragen 4 % bis 71 % des insgesamt assimilierten N. Bezogen auf den unterirdischen N (Wurzeln + Rhizodeposition) beträgt der Anteil der Rhizodeposition zwischen 15 % und 95 %, wobei Leguminosen mit durchschnittlich 73 % einen höheren Anteil als Getreide mit 57 % aufweisen (Wichern et al. 2008).
Die Rhizodeposition wird von den Bodenmikroorganismen schnell als Nahrung aufgenommen und somit zunächst im Boden mikrobiell immobilisiert. Ein Teil des so immobilisierten N wird im Verlauf von Wochen und Monaten aber wieder remineralisiert und steht den Pflanzen wiederum als Nahrungsquelle zur Verfügung. Man nennt diesen Prozess «Immobilisierungs-Mineralisierungs-Umsatz» (Abb. 1). Ein anderer Teil, ca. 50 % des immobilisierten N, wird direkt im Boden als OBS stabilisiert und dient dem Erhalt oder Aufbau des N-Vorrats der OBS (Kallenbach et al. 2016). Die Transformation von «mikrobiellen Residuen» zu organischer Bodensubstanz ist deshalb ein Schlüsselprozess der Bodenbildung und wesentlicher Motor zum Erhalt der Bodenfruchtbarkeit. Werden oberirdische Pflanzenrückstände in den Boden eingebracht, laufen die Prozesse im Grundsatz gleich ab wie bei Wurzeleinträgen.
Die Dynamik wird sehr stark von der Qualität ober- bzw. unterirdischer Rückstände bestimmt. Dabei steuert das C:N-Verhältnis bzw. das Lignin:N-Verhältnis, ob und in welchem Ausmass N aus den Rückständen initial mineralisiert oder immobilisiert wird bzw. wie lange die Remineralisierungsphase dauert. Vereinfacht lassen sich zwei Kategorien bilden:
- Rückstände mit einem C:N-Verhältnis von ca. 9–23 zeigen keine N-Immobilisierung nach Einbringen in den Boden;
- Aus Rückständen mit einem C:N-Verhältnis von ca. 30–136 wird N immobilisiert und später wieder remineralisiert. Das Ausmass des Prozesses hängt zusätzlich vom Ligningehalt der Rückstände ab (Chen et al. 2014). Die zeitliche Dynamik sowie das Ausmass der Stabilisierung im Boden wird von den Bodentemperaturen, der Bodenfeuchte sowie vom Tongehalt des Bodens beeinflusst. Sie ist deshalb sehr stark standortbezogen und erlaubt keine einfachen allgemeingültigen Aussagen (Abb. 1, Tab.).
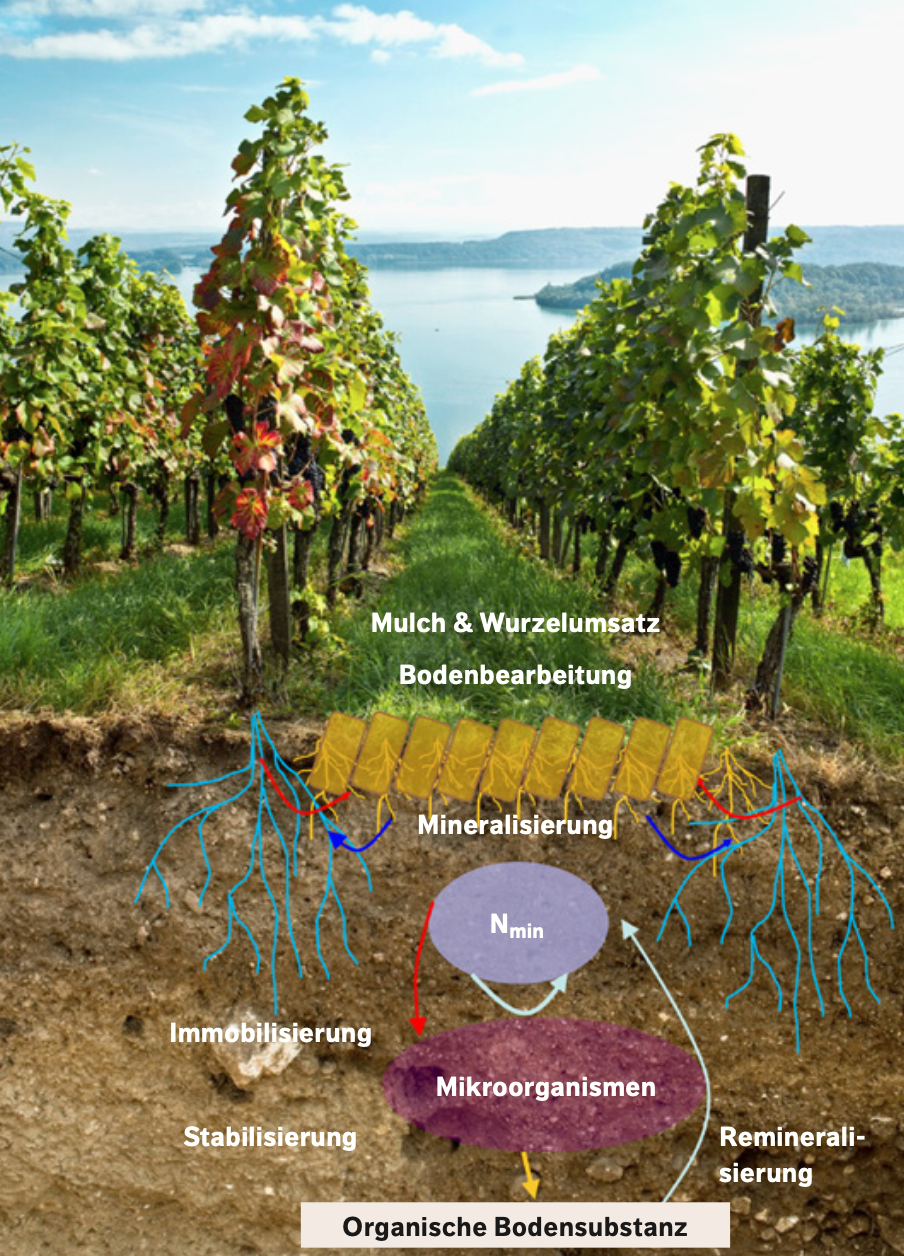 Abb. 1: Schematische Darstellung des Immobilisierungs-Mineralisierungs-Umsatzes. (© Agroscope)
Abb. 1: Schematische Darstellung des Immobilisierungs-Mineralisierungs-Umsatzes. (© Agroscope)
 Abb. 2: Die Grafik zeigt, dass eine frühe N-Mobilisierung für die Rebe keinen Vorteil bringt.
Abb. 2: Die Grafik zeigt, dass eine frühe N-Mobilisierung für die Rebe keinen Vorteil bringt.
Rebberge sind Mischkulturen
Rebanlagen mit spontaner oder gezielt etablierter Begrünung können als Mischkultursystem betrachtet werden. Je nach Anlage und Management der Begrünung kann diese auch zur N-Ernährung der Reben beitragen. Dies wird vor allem in biologischen Systemen genutzt, kann aber in konventionellen Systemen dazu beitragen, den Mineraldüngereinsatz zu reduzieren und weitere Ökosystemleistungen wie Nützlingsförderung, Biodiversität, Reduktion von Nitratauswaschung etc. zu gewährleisten.
Vor allem Gemenge aus Leguminosen und Nichtleguminosen können die N-Ernährung unterstützen. In Studien zu Kleegrasgemengen konnte gezeigt werden, dass die Gräser von dem aus Klee über Rhizodeposition in den Boden eingetragenen N stark profitieren. Rund 40 % des vom Gras im Gemenge aufgenommen N stammt aus dem Transfer vom Klee (Oberson et al. 2013, Hammelehle et al. 2022). Transferraten in dieser Grössenordnung können nur in mehrfach geschnittenen oder gemulchten Systemen erreicht werden, da durch den Schnitt der Wurzelumsatz und damit die N-Rhizodeposition angeregt wird und die sehr konkurrenzfähigen Gräser diesen N effizient aufnehmen können. Die Mechanismen des N-Transfers sind allerdings noch nicht bis ins Detail geklärt. Zu einem gewissen Teil dürften die Reben von den unterirdischen N-Transfers aus Begrünungen profitieren.
Eine bessere N-Nutzungseffizienz lässt sich erzielen, wenn der Mulch nicht auf den Begrünungsstreifen verbleibt, sondern direkt unter die Reben gemulcht wird. Dann wirken die Pflanzenreste wie ein organischer Dünger und die Konkurrenz der Gräser um N in den Begrünungsstreifen ist vermindert.
Immobilisierungsraten
Gemenge mit Leguminosenanteilen haben auch den Vorteil, dass das C:N-Verhältnis der Pflanzenrückstände enger ist und dadurch die Immobilisierungsraten nach einer Bodenbearbeitung (Umbruch der Begrünung) geringer ausfallen. Das vereinfacht, den besten Zeitpunkt für die Bodenbearbeitung zu finden und eine optimierte Synchronisation von N-Bereitstellung (Mineralisierung) aus Pflanzenrückständen und N-Bedarf der Reben zu sichern (Abb. 2). Den exakten Zeitpunkt für die Bodenbearbeitung zu finden, ist relativ schwierig und von vielen Faktoren abhängig wie der Qualität der Begrünung (Alter, C:N, Leguminosenanteil), Intensität der Bodenbearbeitung (Pflügen = Spatenmaschine > Grubber > Eggen), Bodenart (v.a. Tongehalt) und der aktuellen bzw. zu erwartenden Witterung.
Generell sollte auf eine Bodenbearbeitung im Winter verzichtet werden. Bilder wie auf dem Einstiegsbild sollte es besser nicht mehr geben. Als Faustregel kann gelten, die Bodenbearbeitung vor der Hauptwachstumsphase zwischen Austrieb und Vorblüte zu terminieren. So können aus leguminosenreichen Begrünungen zwischen 30 und 80 kg N ha-1 zur Verfügung gestellt werden.
Literatur
Chen B., Liu E., Tian Q. et al., 2014: Soil nitrogen dynamics and crop residues. A review. Agron. Sustain. Dev. 34, 429–442. https://doi.org/10.1007/s13593-014-0207-8
Hammelehle A., Oberson A., Lüscher A., Mäder P., Mayer J., 2018: Above- and belowground nitrogen distribution of a red clover-perennial ryegrass sward along a soil nutrient availability gradient established by organic and conventional cropping systems. Plant Soil 425: 507–525. doi: 10.1007/s11104-018-3559-z.
Hammelehle A., Mayer J., Lüscher A., Mäder P., Oberson A., 2022: 15N natural abundance and 15N leaf labelling methods provide similar estimates of nitrogen transfer from clover to associated grass. In preparation.
Kallenbach C., Frey S. and Grandy A., 2016: Direct evidence for microbial-derived soil organic matter formation and its ecophysiological controls. Nat Commun 7, 13630. https:/doi.org/10.1038/ncomms13630
Oberson A., Frossard E., Bühlmann C., Mayer J., Mäder P., Lüscher A., 2013: Nitrogenfixation and transfer in grass-clover leys under organic and conventional cropping systems. Plant Soil 371: 237–255. doi: 10.1007/s11104-013-1666-4.
Wichern F., Eberhardt E., Mayer J., Joergensen RG., Muller T., 2008: Nitrogen rhizodeposition in agricultural crops: Methods, estimates and future prospects. Soil Biol Biochem 40, 30–48.